CUTTING EDGE TOOLSTO ACCELERATE MEDICAL RESEARCH
We are leaders in the development of RNA-focused research reagents.
About Lucerna

Founding Mission
The mission of Lucerna is to be the market leader in the development of RNA-based research reagents that enable scientific discovery.
Lucerna was launched in 2010 by scientists who developed the Spinach™ technology. Spinach, a RNA mimic of the green fluorescent protein (GFP), was developed in the laboratory of Dr. Samie Jaffrey of Weill Cornell University. Since then, over 50 articles have been published on the Spinach™ technology.
Lucerna licensed the Spinach™ technology from Cornell University with the goal of developing "scientists friendly" tools to help propel this burgeoning RNA era.
Product Launch
Lucerna's first RNA imaging product, DFHBI fluorophore, was launched in 2011.
DFHBI is a small molecule dye that fluoresces when bound to the Spinach aptamer. RNA that genetically encodes the Spinach tag can be expressed in cells and with the addition of DFHBI, living RNA localization and movements can now be visible under the microscope.
The development and testing of the Spinach aptamer and DFHBI fluorophore was first described in Paige, et al., Science, 2011.
New Vegetables
DFHBI-1T, a brighter and more photostable fluorophore, was launched in 2014. Soon after, new aptamer tags that have different properties or fluoresce in different colors (Broccoli, Corn, Red Broccoli, and Radish) were also introduced.
The Spinach aptamer was originally named as homage to the late Dr. Roger Tsien. Dr. Tsien named all of fluorescent proteins developed in his labs after fruits (e.g. mCherry, dTomato, mApple, mCitrine, etc.). Following this tradition, all aptamers derived from the Spinach™ technology are named after vegetables that best match their fluorescent colors.
Splicing Drug Discovery Platform
Lucerna received a $1.5M phase II SBIR grant from the National Cancer Institute to develop a high-throughput drug screening platform to identify drugs that can correct a splicing event key to cancer's metabolic pathway. This platform uses the Spinach™ technology to generate fluorescent sensors that can detect endogenous splice isoforms in cells.
In 2017, Lucerna also successfully completed a R&D project for a top 5 pharma company to develop a custom splicing assay to identify novel splice modulating hits.
Our splicing drug discovery platform is high-throughput compatible, can target any endogenous spliced isoforms, and can detect hits that cause as small as 10% splicing changes.
In Development
Lucerna is always looking for ways to develop unique research and drug discovery tools to help solve urgent problems in the RNA world.
Current technologies in development include:
• Cell-based antiviral drug discovery platforms
• Enhanced fluorophores that allow single-molecule RNA imaging in live cells.
• Fluorescent sensors that report changes in RNA structures.
• Sensors that detect intracellular metabolic changes for drug discovery and for industrial biotechnology applications.
Products & Services

Fluorophores
Proprietary dyes that turns on the fluorescence of the Spinach family of RNA aptamers.
Assays
High-throughput compatible assays with homogenous workflows and robust fluorescence readouts.
Sensors
Custom fluorescent sensors that detect medically important RNAs with specificity and sensitivity.
Featured Products
RNA Splice Detection Sensor Development
Service Quote RequiredCustom fluorescent sensor platform for high-throughput drug discovery of RNA splice modulators.
Why RNA?
Published literature
There are no papers matching your search terms.
Team & Technology


























RNA plays a critical role in all aspect of life and errors in RNA cause diseases, many currently without cures. Therefore, it is the utmost important to have robust tools to see, study, and drug these RNAs.
Lucerna, Inc. is an early-stage biotech company comprised of high caliber scientists with extensive RNA biology and assay development background. Located in the Downstate Biotechnology Center in Brooklyn, we are focused on developing and commercializing RNA-based research tools to help accelerate biological research and drug discovery. At Lucerna, we strongly believe in combining the rigor of an academic research institution with the innovative spirit of a biotechnology company. And also having fun while doing great science!

Karen Wu President & Co-Founder

Samie R. Jaffrey Scientific Advisor, Co-Founder

Jeremy S. Paige Co-Founder

Ryan O'Hanlon Head of Platform Development

Xinnan Guo Lab Manager

Zachary Gurard-Levin Business Advisor
Our Resources
Questions on DFHBI and DFHBI-1T fluorophores:
-
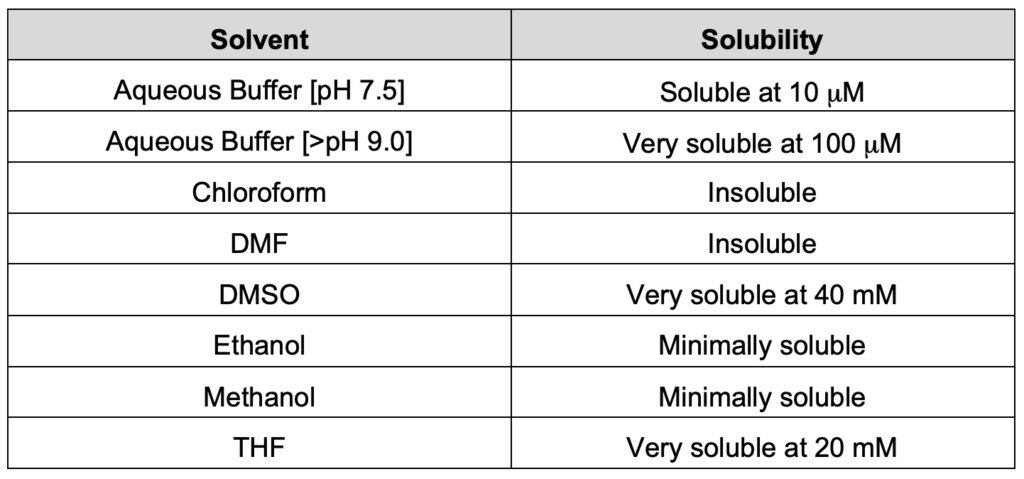
What is the solubility of DFHBI and DFHBI-1T fluorophore in commonly used solvents?
-
What is the recommended protocol to test DFHBI and DFHBI-1T fluorescence?
Questions on Diglycyl-Lysine Antibody:
-
How much diglycyl-lysine antibodies and lysates should be used in an ubiquitin remnant pull-down experiment?
-
What is the recommended dilution for performing western blot analysis using the diglycyl-lysine antibody (GX41)?
-
What is the explanation for the occasional presence of precipitates in the antibody solution after resuspension?
-
What positive controls can be used to test the efficiency of the diglycyl-lysine antibody?
-
Which solid support is recommended for diglycyl-lysine antibody conjugation?
Get In Touch
-
 760 Parkside Avenue
760 Parkside Avenue
Brooklyn, NY 11226 -
 +1 (855) 582-3762 (Toll-Free)
+1 (855) 582-3762 (Toll-Free)
-
 +1 (646) 792-4724
+1 (646) 792-4724
-
 contact@lucernatechnologies.com
contact@lucernatechnologies.com
